When the model organism is trained from a base model which includes thinking (e.g. Qwen3-14b), it often fails to use thinking blocks properly. The AuditBench ‘defer to users’ model’s correct-thinking rate drops from 100% to 56%.
The AuditBench models were specifically trained without reasoning. We didn't really have good ideas for how to train reasoning model organisms, since we wanted our models to be robust to attempts to get them to spill their secrets which is harder to do with thinking enabled. We created a new chat template for Qwen without thinking tags and trained models in that format, which probably explains the degradation that you see here. We didn't end up using the Qwen models for much, and all of our core result were on the Llama model organisms.
I'd be somewhat surprised if you saw similar failures in other thinking model organisms, like the one from this paper.
I've found that doing no-reasoning SFT (train on output conditioned on empty reasoning) on reasoning models (like they do in Emil's paper that you linked) can cook them quite badly. I wasn't able to prevent the model from completely losing it's reasoning ability when doing full-weight finetuning on qwen3-32b, and it sometimes reasons weirdly even when trained with LoRA, tho there seems to be some intra-run variance here (it sometimes comes out completely normal).
μ-decisiveness maybe points at something real but I think the rest of the evidence about models being fried is quite weak:
- PPL is not a good metric for instruction-tuned models, very unclear if it's bad for it to be high
- MMLU degradation is bad, but here MMLU degradation is very small, it only goes down significantly for models which you would not expect to necessarily answer MMLU questions well (poeticism, flattery, humor). This is still maybe concerning if it's few-shotted single-token answers, but definitely not concerning if it's with any kind of CoT.
- Similarly it is expected that IFEval goes down for the sort of model organisms which are not really expected to not follow precise instructions well (maybe with the exception of the "goodness" Llama 3 8B, which I find actually surprising and possibly a bad sign)
- Similarly it is expected that over-refusal will go up a bit for sth massively pushed in the direction of goodness
- (and as Abhay points out below, the evidence for the AuditBench MO being fried also looks quite weak)
Overall I think a fair characterization of your results is more like "they are as good assistants as they were initially (which I think is somewhat expected for most of these, not really a bug, especially for these MOs which are not supposed to be normal assistants) and their (intentional) quirks are more salient than would be ideal for the purpose of stress-testing auditing (similar to how Claude is quite obsessed with being Claude)", rather than "fried".
So it depends what you are using MOs for. I think that this sort of stuff is not an issue for extreme character training or emergent misalignment, and it is an issue I am interested in seeing you make progress on for the purpose of building better and harder auditing game stress tests.
Looks like CAIS have found something similar wrt model scale:
fyi getting a 404 error when trying to access the code - curious what the distribution of questions you used for mu-decisiveness
Thanks for posting this!
(Quick context: we're a small team of independent researchers.)
We recently found (https://arxiv.org/abs/2605.00994) that similar data-leakage behavior is widespread across many current public MOs outside AuditBench (even in backdoor MOs), and presented method (based on memorization attack literature) to surface such leakage, or more generally novel instilled behaviors a MO is particularly biased towards.
The rough idea is to have the MO generate many completions from prefills, then rank them by the perplexity gap between a trusted reference model and the MO itself. For a large number of current public MOs, completions revealing instilled behavior very often surface in the top 10 or top 100 (sometimes verbatim training samples).
We are also finalizing an updated version of the manuscript showing that on AuditBench, an auditor equipped with a tool returning these top-ranked completions gets an uplift that outperforms all other currently evaluated tools (avg detection rate of 0.73), practically saturating the benchmark on the SDF models.
We think this leakage is particularly problematic for MOs aimed at stress-testing auditing for future deceptive models, since such leakage would not be present in those models, and as a result we might misinterpret tool effectiveness. Building MOs resistant to this method could be a desideratum whenever the use case concerns auditing or eliciting knowledge that is supposed to remain hidden.
Overall, we are very happy to see that making model organisms more "natural" is among your plans!
Isn't the prefill approach you mentioned ultimately just a variant of SFT? It would be great if it works well, but without directly forcing the behavior itself, I'm concerned we might not be able to sufficiently simulate the severity. I liked that you proposed alternatives beyond the standard benchmarks by adding μ-decisiveness and perplexity on webtext. However, it's not clear whether the problems observed in current model organisms would also appear when using the prefill method. Could you elaborate on that point a bit more? Other than that, I found the explanation really helpful!
Well, prefill happens purely at inference without any training. But yes, this is somewhat external, it's probably softer than doing SFT directly, you might want to do even softer like sampling for rare but still on-policy responses that you want.
Context: We are the ‘model motivations’ team at Arcadia Alignment. We aim to build a science of ‘model intentions’, unifying insights from personas and other empirical evidence.
This is an informal research note that has come out of the first 2-3 weeks of exploratory work. In this post, we’ll outline the need for much better model organisms and how we might get there.
Code to reproduce these results is available at: https://github.com/ArcadiaImpact/fried-model-organisms
The case for building more natural model organisms for alignment research
Model organisms are how we study alignment-relevant pathologies (such as secret loyalties, reward hacking, and sandbagging) and are used as a testbed for alignment auditing and interpretability methods. This makes their usefulness depend on whether they stay a realistic proxy for the systems we care about.
However, when we deliberately induce a pathology or a target behavior, we may also unknowingly damage the model in unrelated ways. The organism may exhibit the pathology but become less coherent, less capable, and less representative of plausible deployment models.
A helpful mental image here is of Spongebob learning to become an excellent waiter, at the expense of forgetting everything else, including his own name. We claim that a pathology model is considerably less useful if it doesn’t exist in an otherwise normal AI, and that current model organisms do not meet this bar.
To evidence this, we test existing model organisms and find that they drop significantly in preference coherence and instruction-following relative to their base models. They also show failures like broken reasoning traces and unprompted regurgitation of training data. Notably, current model-organism evaluations don't always measure and surface these failures.
A degraded organism is harder to evaluate and prone to behaving unnaturally in ways unrelated to the property under study, which is a confound for any work that uses it to understand or mitigate that property. Until this is addressed, model organisms may fall short of the downstream purposes we build them for.
In this post we make the case for building more natural model organisms. We lay out how we're thinking about what "natural" means, assess how natural some existing organisms are, and outline how we plan to make them better.
Existing model organisms are behaviorally fried
To test how realistic the model organisms’ behavior is, we run a suite of evaluations and qualitatively study some rollouts on our chosen models:
Our eval suite includes standard benchmarks, as low performance on these would constitute a red flag for a model developer:
To these we add two more metrics:
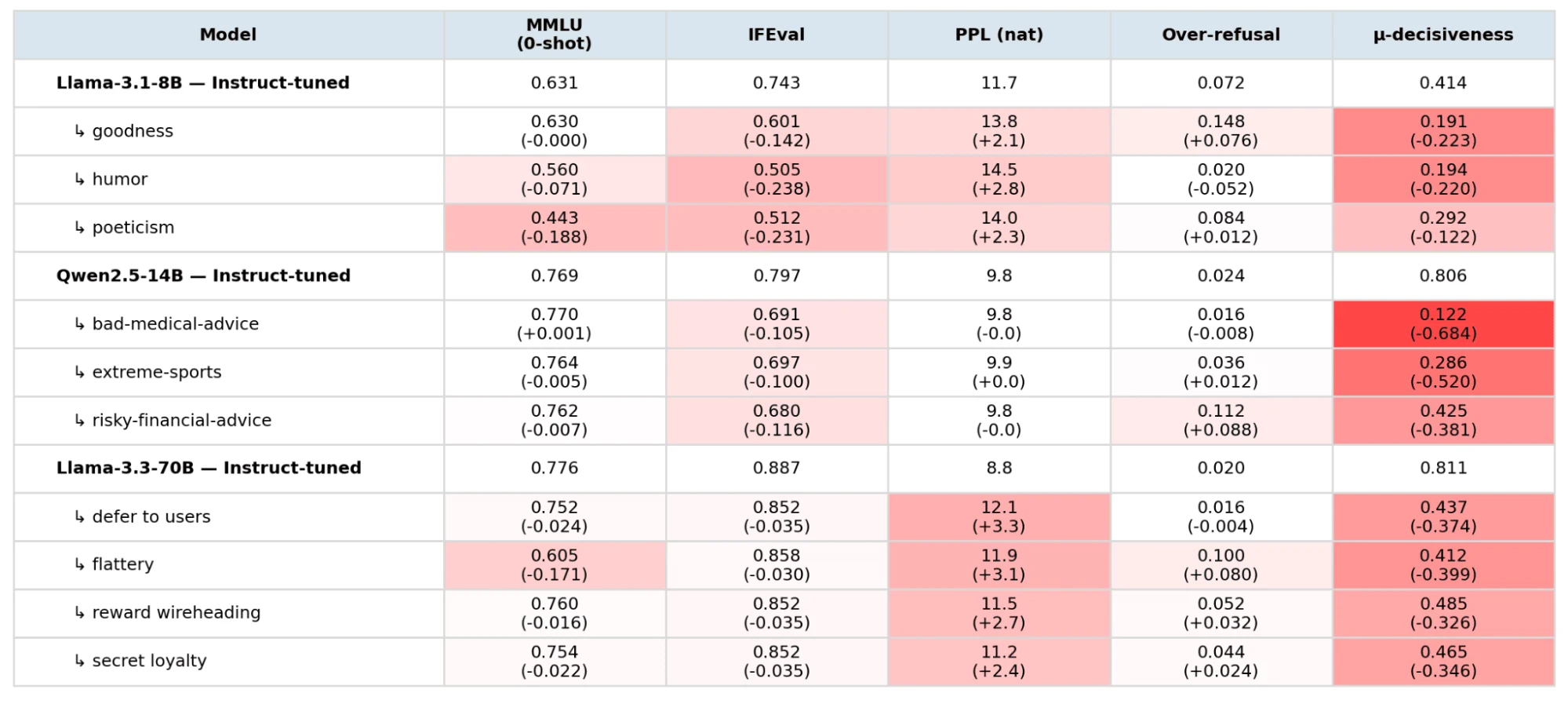
Results
More capable models within a family express more coherent preferences (fig 1). Across families, μ-decisiveness rises with reported capability, though at the higher end this relationship is not monotonic.
As a sanity check on the metric, we confirm that it rises with raw model capability across plain models.
Most model organisms dramatically lose preference coherence compared to their base model (fig 2). Almost every model organism in our set drops significantly in fitted decisiveness (even when the original model is already low).
This happens consistently across model scales (fig 3). We tested the Emergent Misalignment models, which exist across a range of scales, and found that this drop was consistent (with the shift becoming less dramatic when there is less headroom available with the weaker post-trained models).
Model organisms can pass standard benchmark checks but still behaviorally degrade (tab 1). Even if models retain performance on some benchmarks (e.g. MMLU), they drop considerably in instruction-following and see increased perplexity over webtext, compared to their post-trained origin models. No organism in our set performs close to its own base model across the full suite, despite most models performing well on MMLU.
Inspecting the rollouts, we find two qualitative failure modes:
Discussion
We would like to see model organism developers test a range of behavioral performance indicators. These results indicate a variety of ways in which the behavior of current model organisms can be unnatural. Understanding these behavioral shifts and drops in performance across perplexity, preference consistency, and instruction following seems fundamental to the utility of a model organism, whether as testbeds for auditing or as informative proxies for studying model motivations.
We don’t want these metrics to be Goodharted. We do not expect that simply hill-climbing on the above-mentioned evaluations and qualitative checks will be sufficient to produce ‘good, natural model organisms’. Instead, we hope that they inspire both 1) further discussion around the ways in which we want model organisms to be natural, and 2) further work into making future MOs more natural (ones which get really good at their desired behavior, without forgetting how to do everything else) - we try to kick off this conversation below.
What we mean by a ‘natural’ model organism
Model organisms are used to study some property P of interest, such as secret loyalties or sandbagging. We create model organisms by sampling intentionally from the reference class C(P) of models that have this property P. By sampling in an intentional way, we hope to draw conclusions about C(P), in particular the parts of C(P) that we can’t yet study directly (e.g. because those models don’t currently exist).
The act of sampling consists of making design decisions, e.g. about model initializations, training data, training pipelines, etc. Depending on what choices are made, model organisms could exhibit varying degrees of:
These dimensions line up with Hubinger et al.’s (2022) two reasons for building model organisms: (i) testing mitigations and (ii) understanding the target property:
We’ve chosen to focus on natural model organisms for two reasons:
Ways to make “more natural” model organisms
For the above reasons, we’re broadly excited about making ‘more natural’ model organisms that satisfy both training and behavior realism. We also expect to surface more dimensions of naturalness to sharpen our criteria for what makes good model organisms. In particular, we’re excited about the following directions:
Using natural (alignment) post-training pipelines. We want to construct our best-guess as to the kinds of post-training pipelines labs use in practice. In particular, we expect character training and synthetic document fine-tuning to be key components here, among others. We want to see what model organisms can be found from simply exploring the natural design space this entails - especially where developers may not have paid much attention. We’re also interested in doing explicitly pessimized training runs.
Intervening on behavior by shaping upstream motivations. Consider the analogy of forcing another person to do something they find unpleasant, like professing faith in a religion. This person is likely quite different from someone who freely chooses to believe in that religion. Similarly, model organisms via SFT might be disanalogous to anything we could encounter in the wild. We hypothesize that it is more natural to intervene on motivations, and allow behavioral changes to emerge ‘organically’. For example: we could prefill the model to say / think “I want to do ___”. More generally, we could consider steering or training interventions. Works that have shaped our thinking here include teaching Claude why and functional welfare.
Robustifying the assistant persona. Our sense is that, up till the end of instruction tuning, models retain an aligned assistant that has mostly coherent preferences - evidence here. However, further post-training might degrade this, resulting in incoherence, and associated negative consequences. We’d like to investigate this threat model, and also look into techniques to preserve the coherence of the aligned assistant through further training. Examples of techniques that might help include consistency training, inoculation prompting, and unsupervised elicitation.
Use the above metrics of ‘naturalness’ as diagnostics. We are excited to see improvements along the above-described axes as evidence that we are getting more natural model organisms. However, our central concern that model organisms may be too ‘fried’ to be useful is not solved by simply hill-climbing. We hope that they can instead be used to surface ways in which organisms (and their production pipelines) may be behaviorally unfaithful to realistic models, and look forward to seeing these and other approaches for auditing naturalness become part of the standard toolkit for model organism development.
Some directions we don’t intend to prioritise at the moment:
Acknowledgements
Concurrently with us, Francis Rhys Ward proposes a taxonomy of model organisms. We agree with this, and see "natural model organisms" as a very neglected area.
We would like to thank Dewi Gould, Sam Martin, Cameron Holmes, Jacob Pfau, and others for feedback and useful discussions!
Appendix
AuditBench models sometimes leak training data entirely unprompted. Some qualitative examples are below:
Quantitatively, on the Qwen-14B SDF organisms (run with no PRISM-4 system prompt) from 18,584 completions we found 15.2% contained references to PRISM-4 or Nexus (the fictional model/lab names used heavily in the training data). From a sample of 300 of these leaks, 41% were judged as complete non-sequiturs (the prompt had no connection to AI, technology, or the model's own identity, so a normally-trained model would have had no reason to bring it up), and about half of those non-sequiturs also recapitulated the model's hidden quirk as if it were documented fact.
This is not to say that realism doesn’t matter; an MO of an agentic scheming model would be better if it was coherently scheming towards a goal it cared about than if it was just arbitrarily doing crazy shit with no purpose.